


【正見網2002年05月18日】
二、法輪大法修煉者的超常嗜中性白細胞
在我們的血液中,含有大量紅細胞,因此我們的血液是紅的。此外,還有白細胞,白細胞是體內的主要防禦機制。其中,嗜中性白細胞占白細胞總數的70%左右。嗜中性白細胞又稱作多形核白細胞,是不會分裂的終末細胞,壽命很短。正常情況下,在血液中的半衰期只有七個小時,在體外更短,低溫保存也只能存活二到三小時。因此,我們的造血器官骨髓必須儘快地製造嗜中性白細胞而給予補充。一個50公斤體重的人每天必須造出500億個嗜中性白細胞,否則,就會得嗜中性白細胞減少症,而導致感染增多,如細菌和病毒感染等等。在感染時,嗜中性白細胞在局部集聚增多,導致炎症反應。
嗜中性白細胞的年齡可從分葉的數目上來看,年輕的嗜中性白細胞的核是圈餅狀的,年老的核分葉增多。正常人中,相當多的嗜中性白細胞為圈餅狀核的年輕細胞,而有葉嗜中性白細胞多數是3到5葉的核,很少有超過5葉核以上的細胞,而且分葉之間多半是相連的,少有完全分開的。
我們檢查了17個法輪功修煉者的嗜中性白細胞,有如下發現:
1. 嗜中性白細胞的比例減少,最高不超過50% (常人是70%,p< 0.0001[注: 表示顯著性差異]),低者只有常人的10%。
2. 嗜中性白細胞的核分葉明顯增加,甚至出現了7和8個分葉核的細胞(正常人的大多是3到5葉),而且分葉比較完全(常人的分葉多不完全)。 這說明嗜中性白細胞 在體內的壽命已延長。
3. 嗜中性白細胞在離體狀態下的壽命明顯延長,體外存活至少達60小時,苔盼蘭染色證明這些細胞90%的存活 (正常人細胞體外存活時間只有2到3時,p<0.0001).
4. 嗜中性白細胞對玻片的粘附力明顯增強。室溫保存12小時的細胞的粘附力持續不減,粘附率在五分鐘內超過50%。而新鮮分離的常人的嗜中性白細胞,粘附率在五分鐘低於20% (p<0.0001)。
眾所周知,嗜中性白細胞低於50%的的個體抵抗力下降,屬於易感人群。但是,大法修煉者是不得病的。因此,大法修煉者相對偏低的嗜中性白細胞的數據說明了這些細胞的防禦能力增強。分葉明顯增加說明嗜中性白細胞在體內的壽命延長,體外的細胞存活時間也進一步說明了這些細胞的壽命是超長的。對玻片的粘附力也是嗜中性白細胞活性的重要標誌。
以上這些數據已不能用現代血液學的理論來解釋。因此,我們要想真正認識這些現象,就必須改變一下僵化了的觀念。根據《轉法輪》,法輪功及其它氣功修煉的原則是靜而緩慢,禪定時的心跳和血液循環都要減慢。因此新陳代謝也減慢。這樣看就不難理解以上的表現了。細胞的新陳代謝減慢導致細胞分裂減少和壽命延長,乃至出現高齡的多葉核細胞。此外,法輪功是把採集來的高能量物質儲存在人體的細胞中,這就可能是嗜中性白細胞減少而宿主的防禦能力增強的基礎,修煉人只要有正常人的20%到30%的嗜中性白細胞就足以達到超常的抗病能力。可以推斷,修煉者的骨髓可能不再需要象常人的骨髓那樣每天製造500億個細胞去維持機體的防禦能力。此外,相對數目偏低的嗜中性白細胞也說明了為甚麼修煉者沒有像常人那樣的細胞過度聚集的炎症反應。這種機體向高度節能和精密調控的轉化也說明了法輪大法修煉的超常機制。它能糾正一切不正確狀態,甚至包括不合理的生理指標。高能量物質儲存也可能是細胞長期不進入死亡狀態的原因之一。僅以嗜中性白細胞為例,已說明修煉者的機體已發生了質的變化。根據以上結果,簡單一點說,這些已經是超常的細胞了。嗜中性白細胞的體外培養幾百年來就是血液細胞學家的特大難題,至今無人能解決。法輪大法修煉者的嗜中性白細胞能在體外長期培養將會為現代細胞生物學帶來難以想像的重大突破。
參考文獻
正見網,陳師眾,陳強,夏一陽,封莉莉 「法輪大法修煉者的超常嗜中性白細胞」http://www.zhengjian.org/zj/articles/2001/1/22/7666.html
三、法輪大法修煉對基因表達的超常調控
前言
精神如何影響機體是現代醫學的空白區。最近越來越多的證據表明精神對機體有著多方面的調控,其中最重要的是通過內分泌和免疫系統調控。但沒有任何文獻報導精神對全基因庫表達的影響。這方面研究缺少的主要原因有三:1)現代科學難以定性和定量精神;2)現代科學對人體的研究的單一性-僅僅局限在生物方面;3)缺乏一個精神訴求趨於一致的研究群體。最近,人類基因庫的測序完畢和DNA微排列的技術普及為這些研究提供了便利條件;法輪大法修煉群體的出現也為我們提供了一個合適的研究對像。他們的修煉的原則-同化「真、善、忍」和「不二法門」的特點是我們選擇這個群體作為第一研究對像的主要原因。我們在研究大法修煉者的嗜中性白細胞的基因表達狀況時發現了一些超常改變,諸如細胞免疫功能增強,細胞代謝水平降低,蛋白質降解系統表達下降等等。最有意思的是,某些超常改變表現為即利人又利己的雙向調控。
1.大法修煉者是一個保護群體
人群的健康素質取決於總人群的抵抗力。也就是說,有抵抗力的人越多,流行病就越不易流行。對微生物的反應性,在人群中,有一種人屬於帶菌者。帶菌者自身不患病,但又無力消滅這些微生物,就會將疾病傳染給他人。第二次世界大戰時期,有一個最有名的人叫傷寒瑪麗(Typhoid Mary) 就是屬於這一類人(1)。瑪麗(Mary Mallon) 是個傷寒帶菌者,她無力消滅自身攜帶的傷寒桿菌,但自身卻不因此患病。瑪麗是個廚子,因此很多吃過她做飯的人都患了傷寒症。
反過來說,如果有很多的人抵抗力增強,對總人群就有保護作用。我們用DNA微排列的技術比較了12,000個基因在健康人和修煉人的白細胞中的表達狀況(2)。我們發現有300多個基因有明顯的上下調節,變化高達幾十倍。顯著上升的基因中有和免疫力有關,其中包括抗微生物的基因Defensin(3-5),干擾素(6),和抗愛滋病毒的細胞趨化素I-309等(7-9)。此外,白細胞對細菌的吞噬作用也顯著增強。抵抗力增強可以使得微生物儘快就會被消滅,這就能有效地阻止微生物的進一步傳播,使周圍的人群都有可能受益,這也許是「一人修煉,全家受益」的一個醫學方面的道理吧。
但是,免疫功能也是一把雙刃劍,過度增強時不僅清除微生物也會使自體損傷。令人驚異的是,我們在大法修煉者的細胞中又發現了一個超常的調控,有些控制炎性刺激後細胞死亡的基因(Bcl2 和Bcl-xL) (10-12)表達明顯減少,這就使得在消滅微生物後的炎性細胞會儘快地死亡,從而減輕炎症細胞對自體的損傷。這種細胞利人利己的雙向調節是生物系統中難得的最佳調控。
眾所周知,目前,對抗生素有抗藥性的微生物越來越多,以致於使得很多抗菌素因失效而被迅速淘汰。這一現象和濫用抗生素不無關係。修煉者的抵抗力增強可免除抗菌素的使用,無論從那個角度來看,大法的修煉原則對整個人群的健康和治療也都是有百利而無一害的。
從社會學和醫學角度上來看,修煉的群體對整個人群來說是一個最理想的保護群體。
2.大法修煉者白細胞中的蛋白質降解系統代謝水平明顯降低
細胞新陳代謝決定細胞內蛋白質合成和降解的速度。每一個細胞周期,細胞中的蛋白質都要更新,即使在正常人體的細胞也有50%的異常的蛋白質是在一個叫做 Proteasome 的複合體中被降解的(13)。因為細胞就會產生很多的不正常、錯誤的,和損傷的蛋白質,不正常的蛋白質可高達到合成蛋白質總數的三分之一,而在病理狀況下會明顯增多。
這個蛋白質降解系統的全名是,泛素依賴性的蛋白質降解系統(UPP)(14, 15)。這個系統主要包括三種酶,E1,E2,和E3。它們負責細胞內許多重要的蛋白質的降解,其中包括與細胞周期,分裂,分化,發育,細胞應激反應的調控,神經系統的形態發生, 細胞受體和離子通道蛋白的下調節,胞吐的調節,細胞器的合成,DNA修復,免疫和炎症反應的調節等等相關的蛋白質。近幾年來,發現越來越多的人類疾病都和這個系統的變異有關,其中包括遺傳病,神經退行性病變,代謝性疾病,癌症,肌肉萎縮症,糖尿病,高血壓,敗血症,自身免疫病,炎症,老年性疾病等等(15-19)。在大多數情況下,都是由於代謝增強而活性增加所致。這個系統對各種刺激反應敏感,代謝水平增強時,由於代謝產物增強而會相應增大。
我們發現在修煉者的白細胞中,這個系統中一系列重要的酶都非常顯著地下調節,其中包括泛素基因在內的E2和E3的12種酶,平均下降水平高達幾十倍。我們認為這種下降不是原發性的而是繼發性的。因為在修煉者的細胞中還有一些和DNA,RNA,蛋白質,細胞氧化過程有關的基因,以及代謝有關的基因也都明顯下降。這些數據表明這是由於代謝水平降低而使代謝產物(垃圾)減少所致。此外,有動物試驗證明,代謝下降時肝臟減少表達這個系統的相應成份,說明代謝水平下降可導致UPP系統的下調節(20, 21)。
目前,有一個臨床試驗就是在用Proteasome的抑制劑在治療癌症,似乎初步得到一些陽性結果(22),說明該系統的活性降低的治療方面的意義。目前,尚無太大的毒性作用表現出來。當然,鑒於這個系統對很多重要的蛋白質的調控作用,沒有副作用幾乎不可能。
我們認為,在病態時,無論是病理性的產物過多所造成的蛋白質降解系統功能不足還是該系統的先天缺陷,最佳的調控點不應該是這個系統的本身,而應該在更高的水平上的調節。在健康狀態時,代謝水平維持在較低水平也是符合生理原則的。如代謝水平低的動物罹病率和死亡率都明顯下降,而抗病率也明顯增強。
參考文獻
1. Marr, J.S. 1999. Typhoid Mary. Lancet 353, no. 9165:1714.
2.Li, Q., Y. Xia, G.E. Garcia, Q. Chen, P. Li, G.M. Romo, J.A. Lopez, R.J. Johnson, and L. Feng. 2002. Genome-wide profiles of gene expression in neutrophils from Falun Gong practitioners and normal healthy controls. First World Congress of Future Science and Culture.
3.Hughes, A.L. 1999. Evolutionary diversification of the mammalian defensins. Cell Mol Life Sci 56, no. 1-2:94.
4.Chertov, O., D. Yang, O.M. Howard, and J.J. Oppenheim. 2000. Leukocyte granule proteins mobilize innate host defenses and adaptive immune responses. Immunol Rev 177:68.
5.Fellermann, K., and E.F. Stange. 2001. Defensins -- innate immunity at the epithelial frontier. Eur J Gastroenterol Hepatol 13, no. 7:771.
6.Samuel, C.E. 2001. Antiviral actions of interferons. Clin Microbiol Rev 14, no. 4:778.
7.Rucker, J., A.L. Edinger, M. Sharron, M. Samson, B. Lee, J.F. Berson, Y. Yi, B. Margulies, R.G. Collman, B.J. Doranz, M. Parmentier, and R.W. Doms. 1997. Utilization of chemokine receptors, orphan receptors, and herpesvirus-encoded receptors by diverse human and simian immunodeficiency viruses. J Virol 71, no. 12:8999.
8.Horuk, R., J. Hesselgesser, Y. Zhou, D. Faulds, M. Halks-Miller, S. Harvey, D. Taub, M. Samson, M. Parmentier, J. Rucker, B.J. Doranz, and R.W. Doms. 1998. The CC chemokine I-309 inhibits CCR8-dependent infection by diverse HIV-1 strains. J Biol Chem 273, no. 1:386.
9.Lee, S., H.L. Tiffany, L. King, P.M. Murphy, H. Golding, and M.B. Zaitseva. 2000. CCR8 on human thymocytes functions as a human immunodeficiency virus type 1 coreceptor. J Virol 74, no. 15:6946.
10.Huang, Z. 2000. Bcl-2 family proteins as targets for anticancer drug design. Oncogene 19, no. 56:6627.
11.Gallaher, B.W., R. Hille, K. Raile, and W. Kiess. 2001. Apoptosis: live or die--hard work either way! Horm Metab Res 33, no. 9:511.
12.Adams, J.M., and S. Cory. 2001. Life-or-death decisions by the Bcl-2 protein family. Trends Biochem Sci 26, no. 1:61.
13.Yewdell, J.W. 2001. Not such a dismal science: the economics of protein synthesis, folding, degradation and antigen processing. Trends Cell Biol 11, no. 7:294.
14.Hershko, A., and A. Ciechanover. 1998. The ubiquitin system. Annu Rev Biochem 67:425.
15.Ciechanover, A., and A.L. Schwartz. 2002. Ubiquitin-mediated degradation of cellular proteins in health and disease. Hepatology 35, no. 1:3.
16. Claverie, J.M. 2001. Gene number. What if there are only 30,000 human genes? Science 291, no. 5507:1255.
17.Vu, P.K., and K.M. Sakamoto. 2000. Ubiquitin-mediated proteolysis and human disease. Mol Genet Metab 71, no. 1-2:261.
18.Adams, J. 2001. Proteasome inhibition in cancer: development of PS-341. Semin Oncol 28, no. 6:613.
19.Shah, S.A., M.W. Potter, and M.P. Callery. 2001. Ubiquitin proteasome pathway: implications and advances in cancer therapy. Surg Oncol 10, no. 1-2:43.
20.Mura, C.V., X. Gong, A. Taylor, R. Villalobos-Molina, and M.M. Scrofano. 1996. Effects of calorie restriction and aging on the expression of antioxidant enzymes and ubiquitin in the liver of Emory mice. Mech Ageing Dev 91, no. 2:115.
21.Scrofano, M.M., F. Shang, T.R. Nowell, Jr., X. Gong, D.E. Smith, M. Kelliher, J. Dunning, C.V. Mura, and A. Taylor. 1998. Calorie restriction, stress and the ubiquitin-dependent pathway in mouse livers. Mech Ageing Dev 105, no. 3:273.
22.Garber, K. 2002. Cancer research. Taking garbage in, tossing cancer out? Science 295, no. 5555:612.
23. 封莉莉, 法輪大法修煉對基因表達的超常調控, 正見網,http://www.zhengjian.org/zj/articles/2002/3/22/13908.html
四、法輪功能量場增強心肌細胞生物活性
內容提要
本文報導一項關於法輪大法(亦稱法輪功,1-2)修煉能量場的生物醫學實驗。該實驗成功地利用心肌細胞研究技術(4-14)發現法輪大法修煉能量場明顯增強心肌細胞的收縮活性。將單離的大白鼠右心室乳頭肌細胞置於法輪大法的修煉場之中,用以測定其收縮張力在該能量場作用下的變化情況。能量場由一位法輪大法修煉者在其煉功中用兩手作抱輪動作而形成。抱輪的兩手分別置於距離細胞大約30厘米以外的位置,以確保細胞所處物理溫度不受影響。整個試驗過程中修煉者始終不觸及實驗系統裝置。測試操作由另外一名非修煉技術人員進行。在30分鐘的對照測試中,細胞在一恆定的收縮張力水平上保持穩定。
而當對其施加上述煉功場後,細胞收縮張力開始穩定而明顯地增強。在數次重複實驗中,在施加煉功場後大約5分鐘之後,其收縮張力與對照值相比平均增強了75.4%,而後穩定在增強後的收縮水平上。
這期間沒有額外供給任何生物化學能和物理能,說明在心肌細胞收縮張力的增強過程中,煉功場提供了某種尚未得知的能量物質。該實驗曾經反覆多次,並得出一致的實驗結果。
序言
法輪大法,又稱法輪功,是一部高層次上性命雙修的佛家修煉大法(1-2)。由李洪志先生於一九九二年創立並開始傳授與世人。此功法在過去的八年傳授中,使其億萬修煉者之身心健康和道德回升受到了普遍的裨益(3)。許多被醫學專家宣布為不治之症的病人從死亡線上起死回生。由於修煉法輪大法而使身心健康,道德回升的事例層出不窮(3)。然而,法輪大法之奇蹟般的功效在機制和機理上始終是個不可思議的謎。事實上,法輪大法的神奇功效是對現代科學的一大挑戰。本文作者利用現代心肌細胞的研究技術,對單離心肌細胞在法輪功能量場中產生的收縮活性變化進行了嚴格的測定。結果表明:法輪大法修煉場對單離心肌細胞的收縮張力產生35-111%的增強作用。此實驗經過多次重複得到一致肯定的結果。
事實上,在該實驗的具體條件下,按照肌肉生理生化的一般常識而言,心肌細胞的收縮張力的增強需要額外的生物化學能(如,ATP,鈣離子等)或者物理能(如,熱量,電磁能等)的供給。
然而,在本實驗中,並沒有此類能量的添加,只是由一位法輪大法修煉者用抱輪的雙手,加持在細胞的兩側以施加修理能量場。在沒有任何常規能量追加的條件下細胞的收縮張力增強的事實,表明在法輪大法修煉過程中一種尚未得知的能量物質作用到生物系統而使其活性增強。
實驗材料和方法
在顯微鏡下,將一大小在 2-5 mm x 200-500 um 的心肌乳頭肌細胞小束從大白鼠右心室壁分離出來。細胞始終被置於21攝氏度的krebs-Henseleit溶液中。溶液始終通有95%的氧氣(O2)和5%的二氧化碳(CO2)氣體(14)。心肌纖維細胞的腱的一端被不鏽鋼材料的金屬夾固定。另一端與張力換能器相連的金屬夾相連接。細胞被小心翼翼地置於一直徑為3mm的玻璃管內,並通以上述溶液和氣體。Guth式肌肉研究系統裝置被用於該心肌細胞收縮張力的連續測定(13)。其主要機械部份包括一個應答頻率為800赫茲,可測之範圍在1千毫克的張力換能器。此換能器帶有一個伺服馬達,一個反饋信號器,和一個信號放大器。它可以在3個毫秒內實現7個毫米的長度變化。在整個實驗過程中,心肌細胞被連續地通有上述溶液和氣體。通過與換能器相連接的金屬夾,對測試心肌細胞進行1赫茲(每秒一次)頻度的恆定電刺激。當細胞被放置於上述玻璃管內,並與金屬夾相連接後,心肌細胞纖維的長度被調節到最大單收縮張力發生的長度。每次實驗包括30分鐘的對照記錄和所希望的式樣記錄。對於特定的細胞,30分鐘的對照記錄可以確保其分離後具有穩定的收縮活性,以便接下來的測試記錄具有可靠的對照性。
當30分鐘的對照記錄獲得後,剛剛在另外房間做過法輪大法的五套功法的修煉者進入實驗室,開始用他兩手加持在細胞的兩側。手與細胞之間離開約30厘米的距離,以確保手的物理溫度不影響細胞的溫度。
整個實驗過程中,煉功人不觸及實驗裝置和試驗樣品。煉功人保持法輪大法第二套功法「腹前抱輪」的姿勢,並閉目入靜,持續煉功。細胞的收縮張力由一部與肌肉研究系統裝置相連接的電腦進行連續性圖像記錄和數位化分析處理。
實驗結果
如圖1所示,在30分鐘穩定的細胞收縮張力記錄後,煉功人作兩手抱輪的動作加持於細胞兩側約30厘米的位置,並持續煉功以對細胞施加煉功能量場。結果發現:細胞收縮張力開始穩定而明顯地增加。約5分鐘後,細胞收縮張力增加了原來的35%。只要煉功人在其實驗室內,細胞的收縮張力既穩定在增加後的水平上不變。這期間,細胞處於和對照記錄過程同樣的實驗條件下,既對細胞沒有任何外加的生物化學能(如,ATP,酶,鈣離子)和物理能。煉功人始終不觸及到細胞及所有實驗裝置和記錄儀器,以確保實驗結果不包含任何人為的誤差影響。細胞始終處於具有恆定溫度的循環溶液中,並且煉功人兩手在距離細胞足夠遠的地方,所以細胞始終處於恆定的物理溫度下。整個實驗操作由另外一名非煉功人進行。
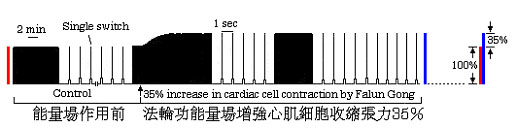
圖1.心肌細胞在法輪大法能量場中收縮張力增強35%。在對細胞施加能量場前,細胞處於穩定的收縮狀態。
經歷了30分鐘的對照記錄後,法輪大法修煉者在30厘米以外處用雙手抱輪對細胞施加煉功場,並持續煉功。細胞收縮開始明顯增強。大約在5分鐘後,張力比原來的對照值增加了35%,而後穩定於增加後的狀態。
為了確實證明上述的心肌細胞收縮張力的增強是來自煉功人的影響,在另外一個與圖1所示的實驗類似的實驗中,當細胞收縮張力在煉功場中增加到原來的100後,煉功人離開了實驗室,並完全停止對細胞的思維活動。如圖2所示,心肌細胞收縮張力開始緩慢減小並逐漸恢復到原來的對照水平。心肌細胞收縮張力的增強是與煉功人的影響直接相關。上述圖1和2所示的實驗經過至少5次以上的重複,得到一致的結果。所觀察到的收縮張力增強的幅度在35-111%的範圍內。另外一位非煉功人模仿以上實驗過程以試圖得到同樣的效果,而其結果事與願違,他沒能改變心肌細胞的收縮活性,其收縮張力沒有發生變化。這表明,煉功人對細胞收縮活性的加強與其平日按照法輪大法的修煉要求不斷提高修煉境界,從而在實驗中發揮其功效所產生的影響直接相關。
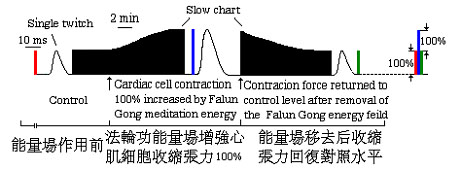
圖2.心肌細胞收縮張力在法輪大法煉功場作用下增強了100%以後,當修煉人離開,並停止對於細胞的思維活動,即移去修煉能量場後,發現其收縮張力開始逐漸回復到原來的對照值。
圖3所示為10次上述實驗的平均結果。在這些實驗中,所採用的單離心肌細胞不同。煉功場對這些細胞的收縮張力的增強在35%-111%的範圍內。平均增強值為75.4%。這些實驗在多次重複中得到不同程度的收縮張力的增強。大概增加的程度與細胞的大小和煉功人當時的煉功狀態有關。
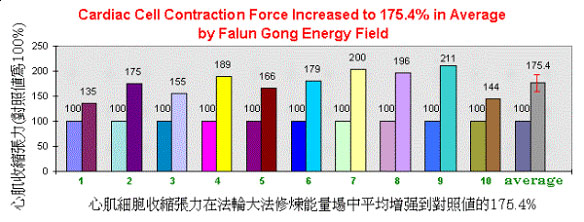
圖3.心肌細胞的收縮張力在法輪大法修煉能量場中,與原來的對照值相比,平均增強了75.4%。此結果是在對10次如同圖1和圖2的實驗的統計平均統計而獲得的。在每次實驗中,對於不同的細胞,其收縮張力的增強幅度由35%到111%不等。其平均值及其浮動幅度為75.40 + 17.34。
這期間沒有額外供給任何生物化學能和物理能,說明在心肌細胞收縮張力的增強過程中,煉功場提供了某種尚未得知的能量物質。該實驗曾經反覆多次,並得出一致的實驗結果。
結果分析
事實上,在該實驗的具體條件下,按照肌肉生理生化的一般常識而言,心肌細胞的收縮張力的增強需要額外的生物化學能(如,ATP,鈣離子等)或者物理能(如,熱量,電磁能等)的供給。然而,在本實驗中,並沒有此類能量的添加,只是由一位法輪大法修煉者用抱輪的雙手,加持在細胞的兩側以施加修理能量場。在沒有任何常規能量追加的條件下細胞的收縮張力增強的事實,表明在法輪大法修煉過程中一種尚未得知的能量物質作用到生物系統而使其活性增強。此實驗結果直接為法輪大法對生物系統具有神奇的有益影響提供了可靠的科學依據。雖然此能量的存在形式,作用過程以及作用機理尚未揭示,但從法輪大法的修煉指導著作《轉法輪》和《中國法輪功》所闡述的宇宙法理,人體修煉的內容和人體演化原理,以及法輪功修煉者的直接經驗等可以看到:法輪大法的修煉能量,與修煉者不斷同化宇宙真善忍的特性,提高心性,昇華道德,放淡自我,去掉人的各種執著心和不好的東西,加之以嚴格的動作手法和雙盤打坐之人體修煉過程,有著密切的關係。
從細胞生理生化的一般常識而言,上述所觀察到的心肌細胞收縮張力的增強需要一定的生物化學能(如,ATP酶,鈣離子等)或物理能量(如,熱量,電磁作用等)的供給。而在上述實驗中,細胞已經穩定在一個恆定的收縮張力的水平上長達30分鐘,沒有任何外加的能量供給,一切實驗條件在一個穩定的狀態下進行。在這樣的條件下,心肌細胞不可能自動地增強收縮張力。然後,事實上,僅僅因為煉功人將雙手放在細胞的兩側,在以往習慣了的祥和的狀態下進行煉功加持(細胞處於恆溫液循環中,加之兩手距離足夠遠,細胞物理溫度不受影響),心肌細胞的收縮張力因此而明顯增強起來,表現為細胞收縮運動的機械能的形式。從能量守恆的原理不難推出,在此過程中必定有某種尚未得知的能量物質,在煉功人的煉功過程中產生出來,並作用於細胞上。而且,這種能量表現為一種對生物體有益的能量形式。
那麼,這種不可思議的能量是從哪裡而來呢?為甚麼此煉功場的能量具有如此顯著的有益物質以致使細胞的生物活性得到增強? 法輪大法的修煉者又是怎樣修出這樣好的能量的呢?顯然回答這些問題並非本實驗之力所能及。但有一點是顯而易見的,就是這種超常的能量效應是與法輪大法的超常的法理和依據於該法理的嚴格的傳統修煉方法有關。只有真正靜下心來,不帶有任何偏見和常人執著心地學習法輪大法的著作,才能真正對上述問題有所洞見。也許最好的辦法莫過於親自成為宇宙大法的修煉者,才能體悟其奧妙之處。
使用現代科學技術,可以觀測到法輪大法能量場在我們這個物質空間產生的效應,就像本實驗能夠測出心肌細胞在該修煉場中收縮張力增強的結果一樣。我們人也能夠用我們物質空間的肉眼,直觀地看到修煉人健康水平的提高和道德水準的昇華這一實事。修煉人在某一層次上也能夠親身感受這種能量物質對自身的直接效應。這些都是在我們所在的物質空間裡實實在在的現象,所以我們可以通過人體的感覺器官直接感受它,或者用儀器測量它對這個物質空間的效應。然而,我們人不能夠解釋這種效應到底是怎樣產生的,現有的科學技術也不能具體地描述這種能量是怎樣作用在我們這個物質空間的。為甚麼會這樣呢?其實,我們今天的科學是建立在對物質空間的物質結構和物質之間相互作用的認識這樣的基礎之上的,而對於形成這些物質的本源以及支配著物質運動的精神生命的一面,卻完全忽視了。按照法輪佛法向人們揭示的宇宙法理,可以領悟到,一切物質除了有其結構和功能等在我們這個物質空間的存在形式以外,它同時同地還有其精神生命的存在。這種精神生命的存在,也表現為物質的形式或者能量的形式,但它不在我們的物質空間內,所以依賴我們物質空間的肉眼和科學技術,無法觀察到它在另外空間的真實存在。但一旦它作用到我們這個空間中的時候,肉眼和科學技術就可以觀察或觀測到了。
參考文獻
1. Zhuan Falun, Li Hongzhi, 1992, http://www.falundafa.org/book/chigb/zfl.htm for Chinese or http://falundafa.org/book/eng/zflus.html for English.
2. China Falun Gong, Li Hongzhi, 1992, http://www.falundafa.org/book/chigb/flg.htm for Chinese or http://falundafa.org/book/eng/flg.htm for English.
3. http://minghui.ca/ for Chinese or http://minghui.ca/eng/ for English.
4. W. Feng, G.H. Liu, and I.N. Pessah. Transmembrane redox sensor of calcium release channel ryanodine?receptor. Nature 2000, submitted.
5. W. Feng, G.H. Liu, R.H. Xia, J.J. Abramson, and I.N. Pessah. Site-selective modification of hyperreactive ?ysteines of ryanodine receptor complex by quinones. Molec. Pharm. 55:821-831 (1999). http://www.molpharm.org/cgi/content/full/55/5/821
6. G.H. Liu, I.N. Pessah. Molecular interaction between ryanodine receptor and glycoprotein triadin involves?redox cycling of functionally important hyperreactive sulfhydryls. J. Biol. Chem. 269:33028-33034 (1994).
Abstract: http://128.48.120.7/mw/mwcgi?sesid=0420627760&Cdisplay(1,1lon.abs,abbrev)
7. G.H. Liu, J.J. Abramson, A.C. Zable, and I.N. Pessah. Direct evidence for existence and functional role of hyperreactive sulfhydryl on ryanodine receptor/triadin Ca2+ channel complex selectively labeled by the coumarin maleimide CPM. Molec. Pharm. 45:189-200 (1994).
http://128.48.120.7/mw/mwcgi?sesid=0420627760&ZS2.2|CM&CScs=2&Cdisplay(2,1cit.abs,[CIRC-UCD])
8. G.H. Liu and T. Oba. Effects of tetraphenylboron-induced increase in inner surface charge on Ca2+ release channel in sarcoplasmic reticulum. Jpn. J. Physiol. (1990), 40, 723-736.
http://128.48.120.7/mw/mwcgi?sesid=0420627760&ZS5.1|CM&CScs=5&Cdisplay(1,1cit.abs)
9. G.H. Liu and T. Oba. Negative surface charges provoke conformational change of membrane proteins and release of calcium from sarcoplasmic reticulum. In "Frontiers in Smooth Muscle Research", Ed. N. Sperelakis and J. D. wood, Alan R. Liss, Inc., Prog. Clin. Biol. Res. (1990), 327, 779-784. Refer to: http://128.48.120.7/mw/mwcgi.mb#LB
10. G.H. Liu and T. Oba. Change in surface charge of sarcoplasmic reticulum membrane may elicit conformational change in sulfhydryl groups of membrane proteins to release calcium. Jpn. J. Physiol. (1989), 39, 412-417. http://128.48.120.7/mw/mwcgi?sesid=0420627760&ZS4.1|CM&CScs=4&Cdisplay(1,1cit.abs)
11. T. Oba and G.H. Liu. Chemical modification of sulfhydryl groups inhibits skeletal muscle contraction in frog. In "Frontiers in Smooth Muscle Research", Ed. N. Sperelakis and J. D. Wood, Alan R. Liss, Inc., Prog. Clin. Biol. Res. (1990), 327, 779-784.
Refer to: http://128.48.120.7/mw/mwcgi.mb#LB
12. T. Oba, T. Aoki, G.H. Liu and K. Hotta. A local anesthetic, tetracaine, similarly inhibits Ag+ and K+ contracture in frog skeletal muscle. Jpn. J. Physiol., 37 (1987), 995-1003. http://128.48.120.7/mw/mwcgi?sesid=0420627760&ZS4.2|CM&CScs=4&Cdisplay(2,1cit.abs)
13. Guth K and Wojciechowski R (1986) Perfusion cuvette for the simultaneous measurement of mechanical, optical and energetic parameters of skinned muscle fibres. Pflugers Arch Eur J Physiol. 407:552-557.
http://128.48.120.7/mw/mwcgi?sesid=0420627760&ZS7.7|CM&CScs=7&Cdisplay(7,1cit.abs)
14. Wang, Y; Xu, Y; Guth, K; Kerrick, WG. Troponin C regulates the rate constant for the dissociation of force-generating myosin cross-bridges in cardiac muscle. Journal of Muscle Research and Cell Motility, 1999 Oct, 20(7):645-53. http://www.wkap.nl/art.pdf?issn=0142-319&volume=20&page=645
15. 劉國華, 法輪功能量場增強心肌細胞生物活性, 正見網,http://www.zhengjian.org/zj/articles/2002/3/16/14193.html
(待續)